
Preventing Extinction of the Critically Endangered Dactylorhiza incarnata subsp. ochroleuca in Britain Using Symbiotic Seedlings for Reintroduction

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
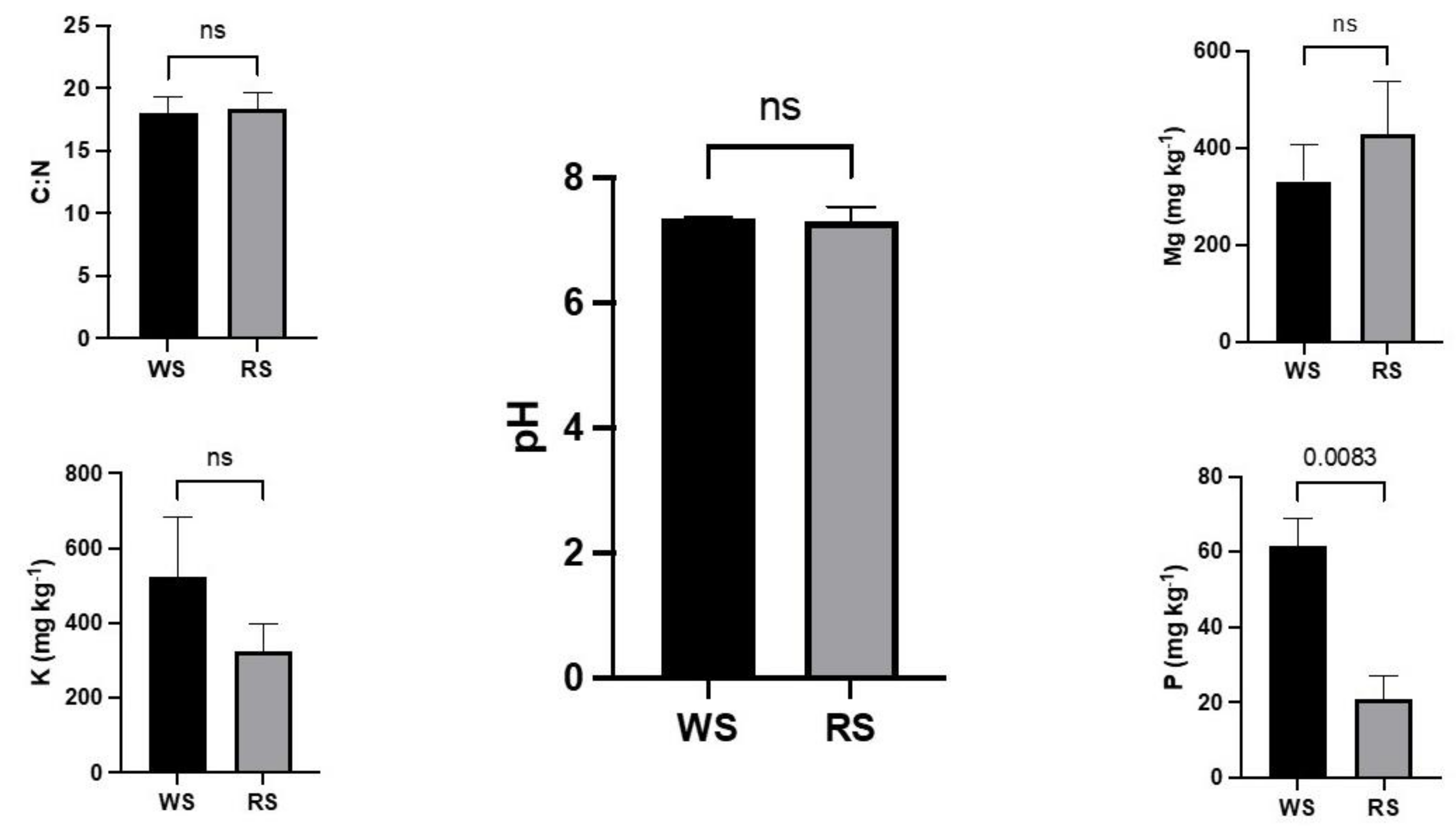
2.1. Soil Analysis
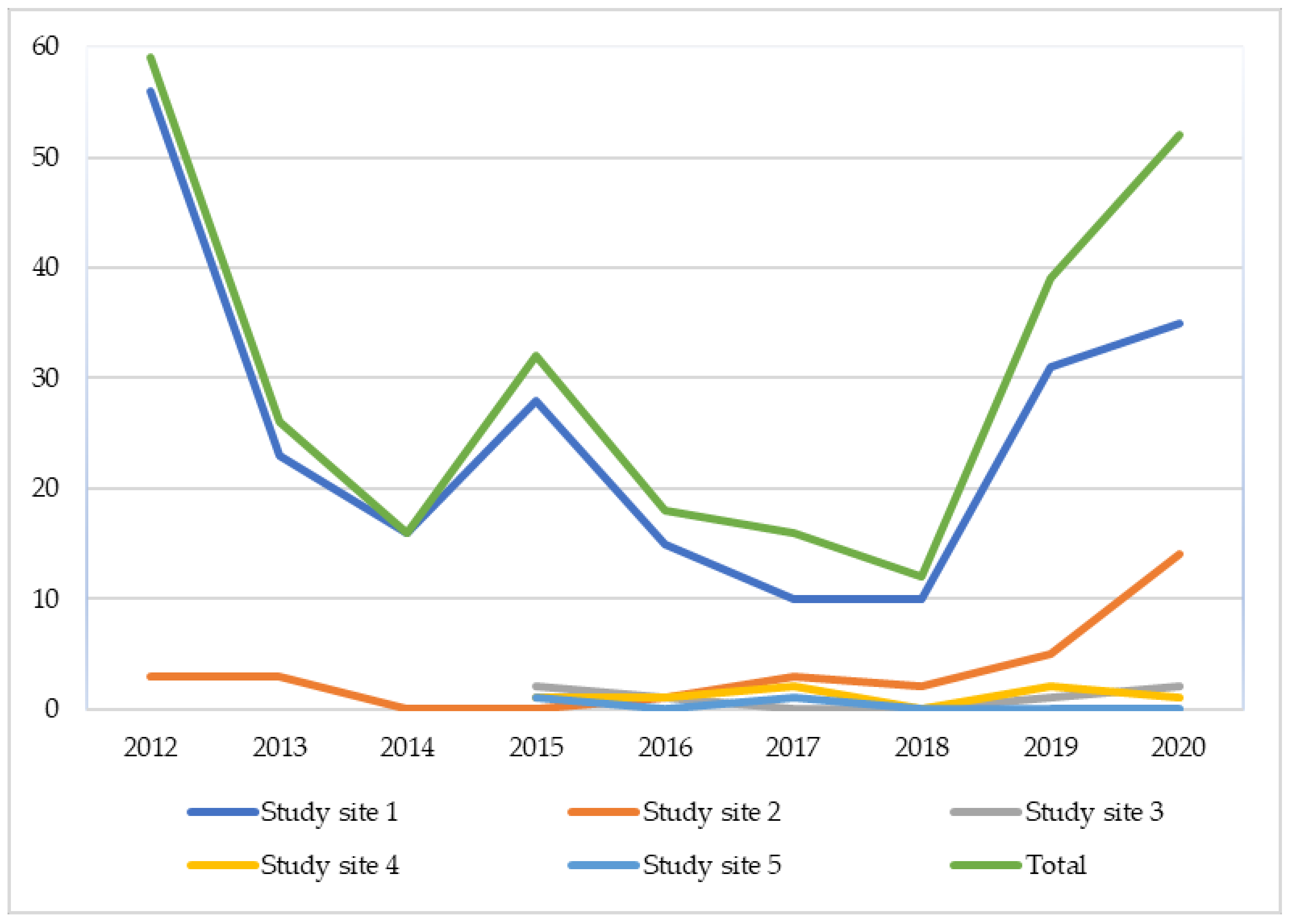
2.2. Population Count and Seed Collecting
2.3. Identification of Compatible Mycorrhizal Fungus for Seed Germination
2.4. In Vitro Seed Germination
2.5. Transfer of Seedlings to Preweaning Medium
2.6. Resilience of Seedlings at Reduced Moisture Content
2.7. Reintroduction of Symbiotic Seedlings
3. Results
3.1. Soil Analysis
3.2. Population Count and Seed Collecting
3.3. Identification of Compatible Mycorrhizal Fungus for Seed Germination
3.4. In Vitro Seed Germination
3.5. Transfer of Seedlings to Preweaning Medium
3.6. Resilience of Seedlings at Reduced Moisture Content
3.7. Reintroduction of Symbiotic Seedlings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reiter, N.; Whitfield, J.; Pollard, G.; Bedggood, W.; Argall, M.; Dixon, K.; Davis, B.; Swarts, N. Orchid re-introductions: An evaluation of success and ecological considerations using key comparative studies from Australia. Plant Ecol. 2016, 217. [Google Scholar] [CrossRef]
- Downing, J.L.; Liu, H.; Shao, S.; Wang, X.; McCormick, M.; Deng, R.; Gao, J. Contrasting changes in biotic interactions of orchid populations subject to conservation introduction vs. conventional translocation in tropical China. Biol. Conserv. 2017, 212. [Google Scholar] [CrossRef]
- Fay, F.M. British and Irish orchids in a changing world. Curtis’s Bot. Mag. 2015, 32, 3–23. [Google Scholar] [CrossRef]
- Bell, J.; Yokoya, K.; Kendon, J.P.; Sarasan, V. Diversity of root-associated culturable fungi of Cephalanthera rubra (Orchidaceae) in relation to soil characteristics. PeerJ 2020, 8. [Google Scholar] [CrossRef] [Green Version]
- Willmer, P. Climate Change: Bees and Orchids Lose Touch. Curr. Biol. 2014, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barman, D.; Devadas, R. Climate change on orchid population and conservation strategies: A review. J. Crop Weed 2013, 9, 1–12. [Google Scholar]
- Kull, T.; Hutchings, M.J. A comparative analysis of decline in the distribution ranges of orchid species in Estonia and the United Kingdom. Biol. Conserv. 2006, 129. [Google Scholar] [CrossRef]
- Bidartondo, M.I.; Duckett, J.G. Conservative ecological and evolutionary patterns in liverwort-fungal symbioses. Proc. Biol. Sci. 2010, 277, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Stroh, P.A.; Leach, S.J.; August, T.A.; Walker, K.J.; Pearman, D.A.; Rumsey, F.J.; Harrower, C.A.; Fay, M.F.; Martin, J.P.; Pankhurst, T.; et al. A Vascular Plant Red List for England; Botanical Society of Britain and Ireland: Bristol, UK, 2014. [Google Scholar]
- Brown, A.E.; Burn, A.J.; Hopkins, J.J.; Way, S.F. The Habitats Directive: Selection of Special Areas of Conservation in the UK: Report No. 270; JNCC: Peterborough, UK, 1997. [Google Scholar]
- Phillips, R.D.; Reiter, N.; Peakall, R. Orchid conservation: From theory to practice. Ann. Bot. 2020, 126. [Google Scholar] [CrossRef] [PubMed]
- Bidartondo, M.I.; Read, D.J. Fungal specificity bottlenecks during orchid germination and development. Mol. Ecol. 2008. [Google Scholar] [CrossRef] [Green Version]
- Jacquemyn, H.; Honnay, O.; Cammue, B.P.A.; Brys, R.; Lievens, B. Low specificity and nested subset structure characterize mycorrhizal associations in five closely related species of the genus Orchis. Mol. Ecol. 2010, 19. [Google Scholar] [CrossRef]
- McCormick, M.K.; Whigham, D.F.; O’Neill, J. Mycorrhizal diversity in photosynthetic terrestrial orchids. New Phytol. 2004, 163. [Google Scholar] [CrossRef] [PubMed]
- Ruibal, M.P.; Triponez, Y.; Smith, L.M.; Peakall, R.; Linde, C.C. Population structure of an orchid mycorrhizal fungus with genus-wide specificity. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Reiter, N.; Lawrie, A.C.; Linde, C.C. Matching symbiotic associations of an endangered orchid to habitat to improve conservation outcomes. Ann. Bot. 2018. [Google Scholar] [CrossRef] [PubMed]
- Yokoya, K.; Zettler, L.W.; Kendon, J.P.; Bidartondo, M.I.; Stice, A.L.; Skarha, S.; Corey, L.L.; Knight, A.C.; Sarasan, V. Preliminary findings on identification of mycorrhizal fungi from diverse orchids in the Central Highlands of Madagascar. Mycorrhiza 2015, 25, 611–625. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2. [Google Scholar] [CrossRef] [PubMed]
- Fageria, N.K.; Baligar, V.C.; Moreira, A.; Moraes, L.A.C. Soil phosphorousiInfluence on growth and nutrition of tropical legume cover crops in acidic soil. Commun. Soil Sci. Plant Anal. 2013, 44, 3340–3364. [Google Scholar] [CrossRef]
- Taylor, D.L.; McCormick, M.K. Internal transcribed spacer primers and sequences for improved characterization of basidiomycetous orchid mycorrhizas. New Phytol. 2008, 177. [Google Scholar] [CrossRef] [PubMed]
- Hosomi, S.T.; Santos, R.B.; Custodio, C.C.; Seaton, P.T.; Marks, T.R.; Machado-Neto, N.B. Preconditioning Cattleya seeds to improve the efficacy of the tetrazolium test for viability. Seed Sci. Technol. 2011, 39. [Google Scholar] [CrossRef]
- Clements, M.A.; Muir, H.; Cribb, P.J. A preliminary report on the symbiotic germination of European terrestrial orchids. Kew Bull. 1986, 41, 437. [Google Scholar] [CrossRef]
- Rafter, M.; Yokoya, K.; Schofield, E.J.; Zettler, L.W.; Sarasan, V. Non-specific symbiotic germination of Cynorkis purpurea (Thouars) Kraezl., a habitat-specific terrestrial orchid from the Central Highlands of Madagascar. Mycorrhiza 2016. [Google Scholar] [CrossRef]
- Arnon, D.I.; Hoagland, D.R. Crop production in artificial culture solutions and in soils with special reference to factors influencing yields and absorption of inorganic nutrients. Soil Sci. 1940, 50, 463–485. [Google Scholar]
- Foley, M.J.Y. Dactylorhiza incarnata (L.) Soó subsp. ochroleuca (Wüstnei ex Boll) P. F. Hunt and Summerh. (Orchidaceae): A comparison of British and European Plants. Watsonia 2000, 23, 299–303. [Google Scholar]
- Albrecht, M.A.; Guerrant, E.O.; Maschinski, J.; Kennedy, K.L. A long-term view of rare plant reintroduction. Biol. Conserv. 2011, 11, 2557–2588. [Google Scholar] [CrossRef]
- Bailarote, B.C.; Lievens, B.; Jacquemyn, H. Does mycorrhizal specificity affect orchid decline and rarity? Am. J. Bot. 2012, 99, 1655–1665. [Google Scholar] [CrossRef] [PubMed]
- Jacquemyn, H.; Deja, A.; de Hert, K.; Bailarote, C.B.; Lievens, B. Variation in mycorrhizal associations with tulasnelloid fungi among populations of five Dactylorhiza species. PLoS ONE 2012, 7, e42212. [Google Scholar] [CrossRef] [Green Version]
- Fuji, M.; Miura, C.; Yamamoto, T.; Komiyama, S.; Suetsugu, K.; Yagame, T.; Yamato, M.; Kaminaka, H. Relative effectiveness of Tulasnella fungal strains in orchid mycorrhizal symbioses between germination and subsequent seedling growth. Symbiosis 2020, 81. [Google Scholar] [CrossRef]
- Yokoya, K.; Jacob, A.S.; Zettler, L.W.; Kendon, J.P.; Menon, M.; Bell, J.; Rajaovelona, L.; Sarasan, V. Fungal diversity of selected habitat specific Cynorkis Species (Orchidaceae) in the Central Highlands of Madagascar. Microorganisms 2021, 9, 792. [Google Scholar] [CrossRef]
- Yokoya, K.; Zettler, L.W.; Bell, J.; Kendon, J.P.; Jacob, A.S.; Schofield, E.; Rajaovelona, L.; Sarasan, V. The Diverse assemblage of fungal endophytes from orchids in Madagascar linked to abiotic factors and seasonality. Diversity 2021, 13, 96. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Waud, M.; Merckx, V.S.; Brys, R.; Tyteca, D.; Hedrén, M.; Lievens, B. Habitat-driven variation in mycorrhizal communities in the terrestrial orchid genus Dactylorhiza. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Mujica, M.I.; Saez, N.; Cisternas, M.; Manzano, M.; Armesto, J.J.; Pérez, F. Relationship between soil nutrients and mycorrhizal associations of two Bipinnula species (Orchidaceae) from central Chile. Ann. Bot. 2016, 118. [Google Scholar] [CrossRef] [Green Version]
- Janissen, B.; French, G.; Selby-Pham, J.; Lawrie, A.C.; Huynh, T. Differences in emergence and flowering in wild, re-introduced and translocated populations of an endangered terrestrial orchid and the influences of climate and orchid mycorrhizal abundance. Aust. J. Bot. 2021, 69. [Google Scholar] [CrossRef]
- Kendon, J.P.; Yokoya, K.; Zettler, L.W.; Jacob, A.S.; McDiarmid, F.; Bidartondo, M.I.; Sarasan, V. Recovery of mycorrhizal fungi from wild collected protocorms of Madagascan endemic orchid Aerangis ellisii (B.S. Williams) Schltr. and their use in seed germination in vitro. Mycorrhiza 2020, 30. [Google Scholar] [CrossRef]
- Huang, H.; Zi, X.-M.; Lin, H.; Gao, J.-Y. Host-specificity of symbiotic mycorrhizal fungi for enhancing seed germination, protocorm formation and seedling development of over-collected medicinal orchid, Dendrobium devonianum. J. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.-Y.; Shao, S.-C.; Liu, S.-J.; Gao, J.-Y. Do the fungi associated with roots of adult plants support seed germination? A case study on Dendrobium exile (Orchidaceae). Glob. Ecol. Conserv. 2019, 17. [Google Scholar] [CrossRef]
- Valadares, R.B.; Pereira, M.C.; Otero, J.T.; Cardoso, E.J. Narrow fungal mycorrhizal diversity in a population of the orchid Coppensia doniana. Biotropica 2012, 44, 114–122. [Google Scholar] [CrossRef]
- Zettler, L.W.; Piskin, K.A. Mycorrhizal fungi from protocorms, seedlings and mature plants of the Eastern Prairie Fringed Orchid, Platanthera leucophaea (Nutt.) Lindley: A Comprehensive List to Augment Conservation. Am. Midl. Nat. 2011. [Google Scholar] [CrossRef]
- Harrap, A.; Harrap, S. Orchids of Britain & Ireland: A Field and Site Guide; A & C Black: London, UK, 2010. [Google Scholar]
- Jasinge, N.U.; Huynh, T.; Lawrie, A.C. Changes in orchid populations and endophytic fungi with rainfall and prescribed burning in Pterostylis revoluta in Victoria, Australia. Ann. Bot. 2018, 121. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Feng, C.L.; Luo, Y.B.; Chen, B.S.; Wang, Z.S.; Gu, H.Y. Potential challenges of climate change to orchid conservation in a Wild Orchid Hotspot in Southwestern China. Bot. Rev. 2010, 76, 174–192. [Google Scholar] [CrossRef]
- Illyés, Z.; Halász, K.; Rudnóy, S.; Ouanphanivanh, N.; Garay, T.; Bratek, Z. Changes in the diversity of the mycorrhizal fungi of orchids as a function of the water supply of the habitat. J. Appl. Bot. Food Qual. 2010, 83, 28–36. [Google Scholar]
- Kohout, P.; Těšitelová, T.; Roy, M.; Vohník, M.; Jersáková, J. A diverse fungal community associated with Pseudorchis albida (Orchidaceae) roots. Fungal Ecol. 2013, 6, 50–64. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Orchid Genotype | Closest Match | %ID | Identity | Isolates |
|---|---|---|---|---|
| Dactylorhiza praetermissa | JX649077 | 96.7 | Tulasnellaceae Tul1 | 03DPR003 03DPR004 03DPR005 03DPR006 03DPR007 03DPR008 03DPR009 03DPR011 |
| Dactylorhiza fuchsii | JX649080 | 93.5 | Tulasnellaceae Tul2 | 01DFU012 01DFU030 01DFU032 01DFU034 01DFU042 01DFU043 01DFU052 01DFU055 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarasan, V.; Pankhurst, T.; Yokoya, K.; Sriskandarajah, S.; McDiarmid, F. Preventing Extinction of the Critically Endangered Dactylorhiza incarnata subsp. ochroleuca in Britain Using Symbiotic Seedlings for Reintroduction. Microorganisms 2021, 9, 1421. https://doi.org/10.3390/microorganisms9071421
Sarasan V, Pankhurst T, Yokoya K, Sriskandarajah S, McDiarmid F. Preventing Extinction of the Critically Endangered Dactylorhiza incarnata subsp. ochroleuca in Britain Using Symbiotic Seedlings for Reintroduction. Microorganisms. 2021; 9(7):1421. https://doi.org/10.3390/microorganisms9071421
Chicago/Turabian StyleSarasan, Viswambharan, Tim Pankhurst, Kazutomo Yokoya, Sridevy Sriskandarajah, and Faye McDiarmid. 2021. "Preventing Extinction of the Critically Endangered Dactylorhiza incarnata subsp. ochroleuca in Britain Using Symbiotic Seedlings for Reintroduction" Microorganisms 9, no. 7: 1421. https://doi.org/10.3390/microorganisms9071421