
Uromys aplini, Cramb & Hocknull & Price, 2020
|
publication ID |
https://doi.org/ 10.3853/j.2201-4349.72.2020.1731 |
|
persistent identifier |
https://treatment.plazi.org/id/2750B75B-FFD7-FFB6-FC03-88987215F87B |
|
treatment provided by |
Felipe |
|
scientific name |
Uromys aplini |
| status |
sp. nov. |
Uromys aplini sp. nov.
urn:lsid:zoobank.org:act:C52317A8-D118-4E10-AA9C-21DF62C8EECA
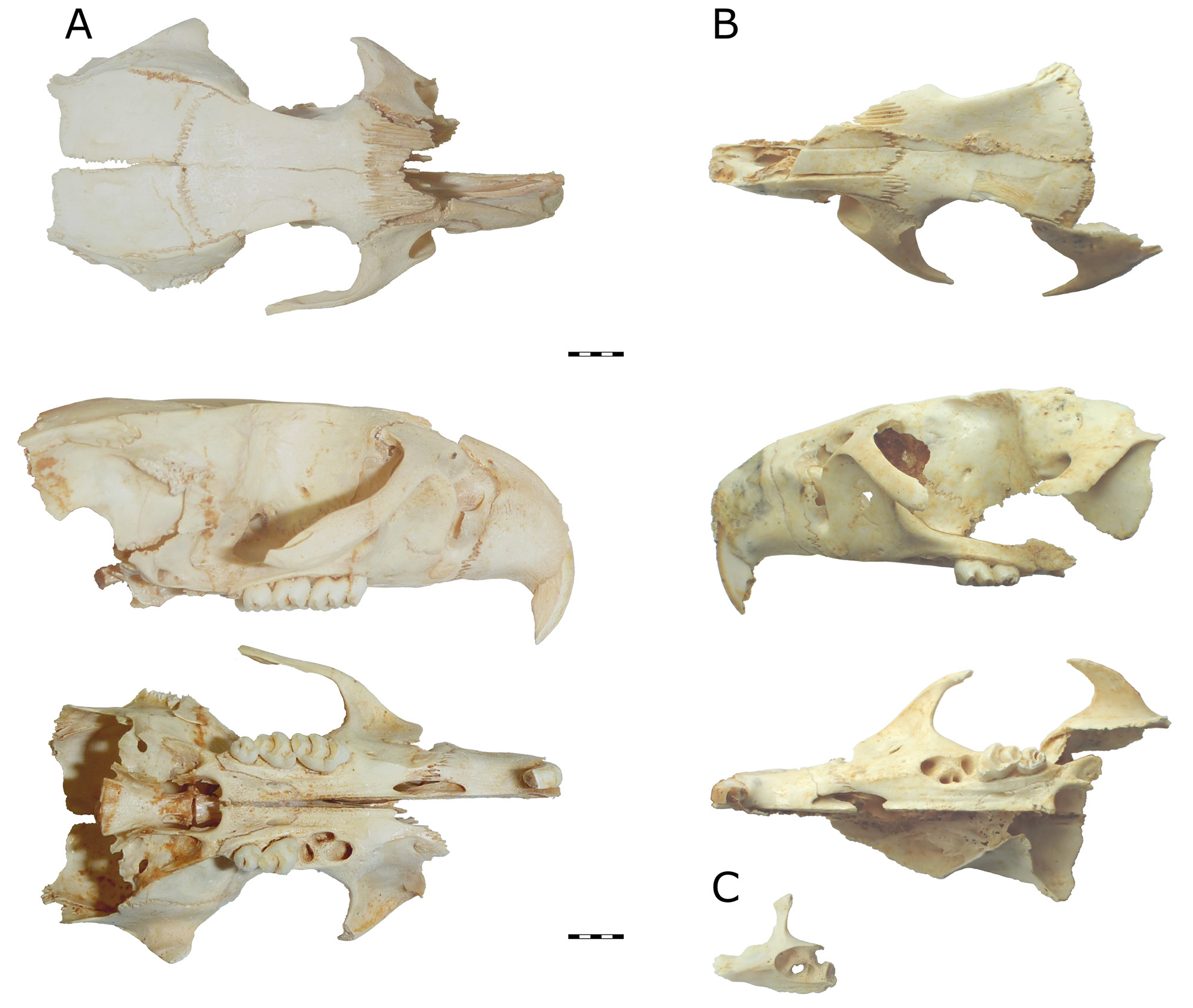
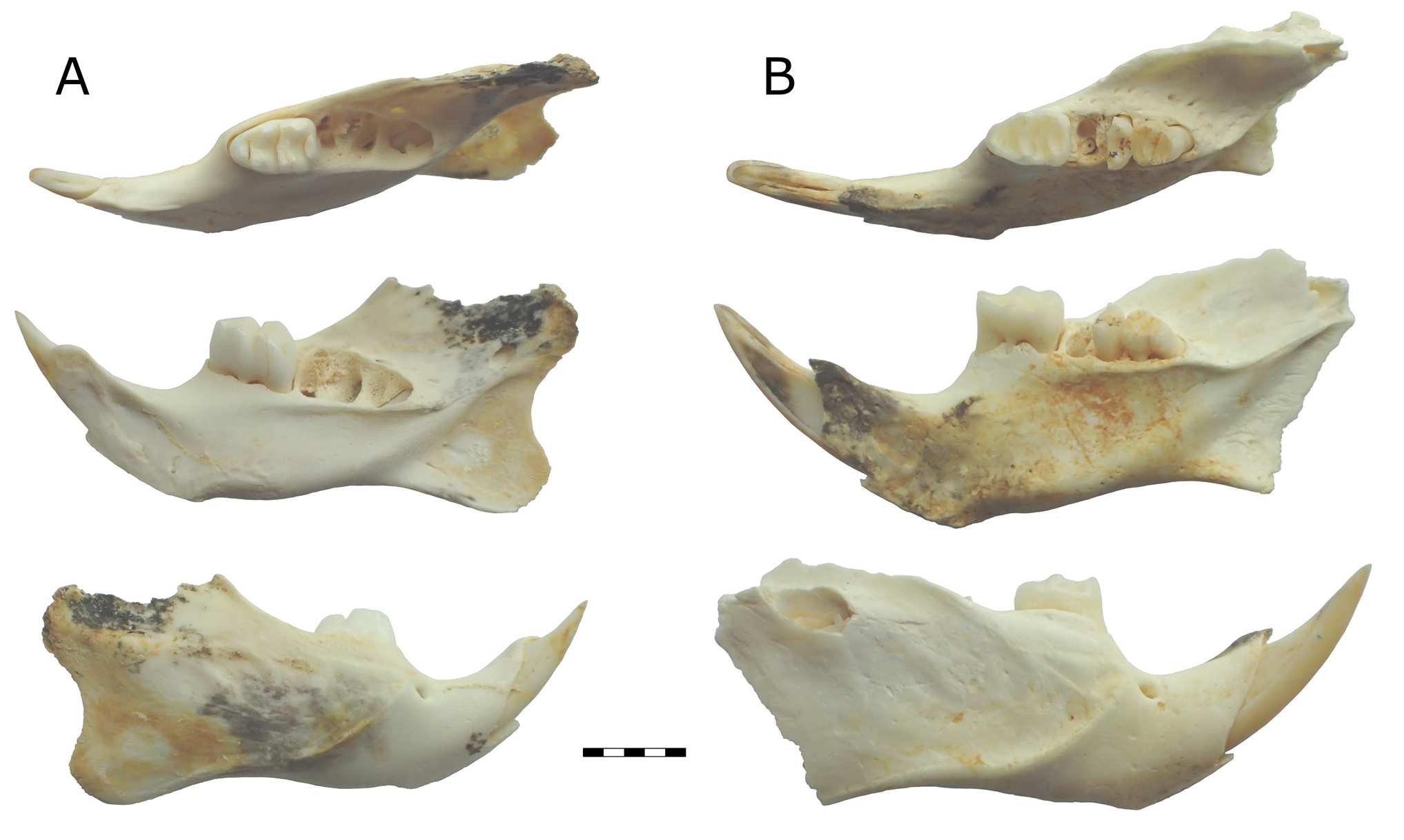
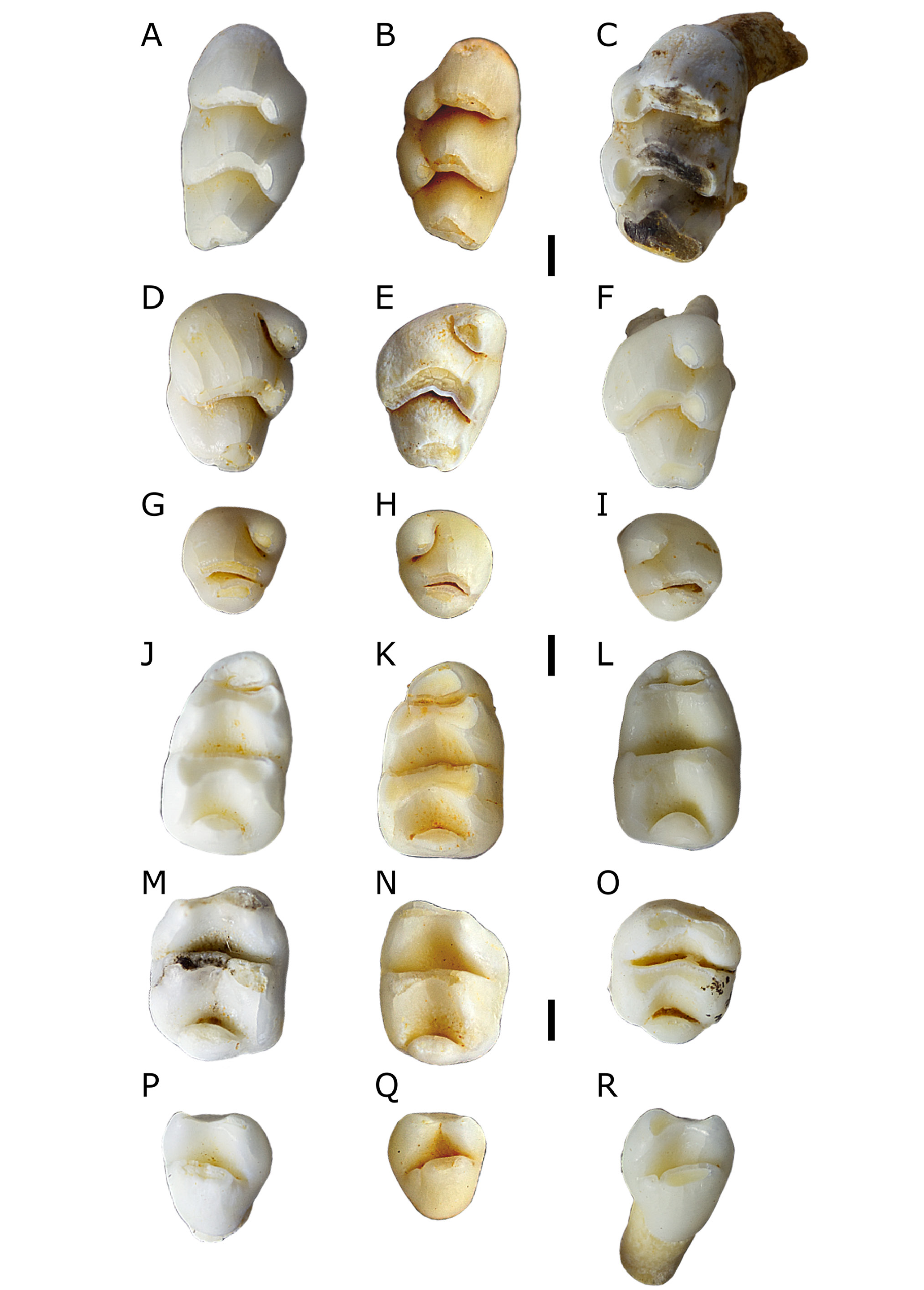
Figs 3C, 3D View Figure 3 , 5–7 View Figure 5 View Figure 6 View Figure 7
Holotype. QMF52014 (Queensland Museum fossil specimen) partial skull, QML1313 (Queensland Museum fossil locality) Speaking Tube Cave , Mount Etna , eastern central Queensland. Deposit has a minimum age of c. 280 ka ( Hocknull et al., 2007) . Paratypes. QMF55753 partial skull ; QMF55542 right mandible with M 1; both specimens have same locality as holotype , QML1313 .
Material examined. QML 1311H: QMF 55547 right M 1, QMF 55548 right M 2, QMF 55549 right M 3, QMF 55550 left M 1, QMF 55551 right M 1, QMF 55552 left M 3, QMF 60125 right M 1; QML 1313: QMF 52014 partial skull, QMF 55522 left M 1, QMF 55523 left M 1, QMF 55524 right M 1, QMF 55525 left M 2, QMF 55526 right M 2, QMF 55527 right M 2, QMF 55528 right M 3, QMF 55529 left M 3, QMF 55530 left M 3, QMF 55531 left M 1, QMF 55532 left M 1, QMF 55533 left M 1, QMF 55534 right M 2, QMF 55535 left M 2, QMF 55536 left M 2, QMF 55537 right M 3, QMF 55538 left M 3, QMF 55539 left M 3, QMF 55540 left I 1, QMF 55541 left maxilla fragment, QMF 55543 right mandible with M 1 and M 3, QMF 55544 right M 3; QML 1313A: QMF 55545 left M 1, QMF 55546 right M1; QML 1312: QMF 55340 left M 1. Additional specimens were also recovered from QML 1284, QML 1284A, QML 1311C/D, QML 1311J, QML 1383, QML 1384 LU, and QML 1385.
Age Range. Chibanian (Middle Pleistocene), chronometrically dated to>500 ka to c. 205 ka.
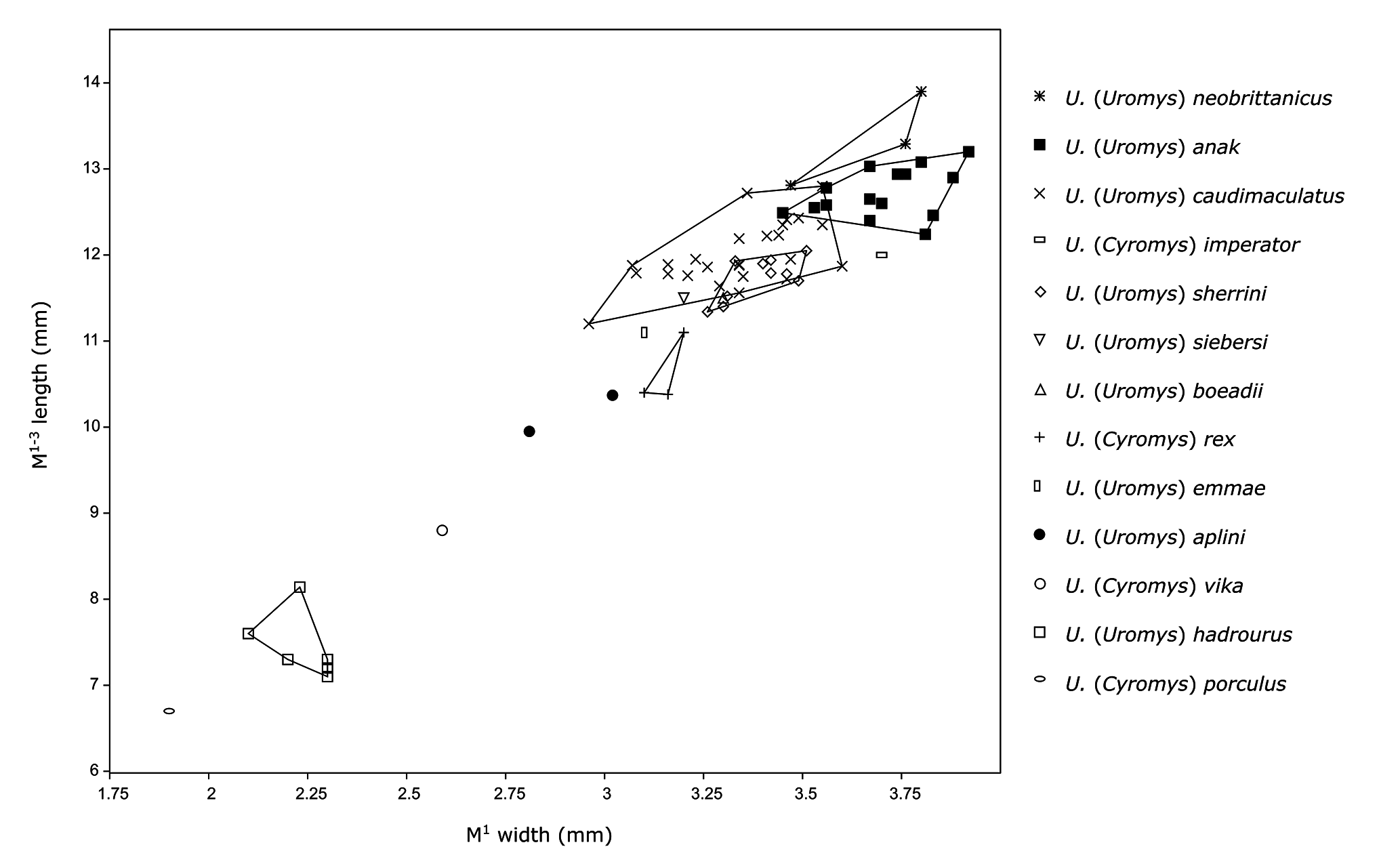
Diagnosis. Large Uromys , but smaller than most species of Uromys (Uromys) with the exception of U. hadrourus ( Fig. 8 View Figure 8 ; Table 2); it is distinguished on the following combination of characters: posterior indent on T8–9 of M 1–2 poorly developed; molar enamel ornament moderately developed; anterior palatal foramina short, shared equally between premaxilla and maxilla; rostrum proportionally short and robust; supraorbital ridges and postorbital processes absent. Features that further distinguish U. aplini from all other species of Uromys are listed in the Remarks section.
Groves & Flannery (1994) divided Uromys into two subgenera: U. ( Uromys ) and U. ( Cyromys ). Uromys aplini is placed in U. ( Uromys ) on the basis of the following diagnostic characters identified by Groves & Flannery (1994): short, slit-like anterior palatal foramina; simplified, elongate molars; reduced M 3 / 3; posteriorly lengthened bony palate; reduced anterior lophid on M 1, which fuses to middle lophid after moderate wear; zygomatic arches swing posteriorly and ventrally to level of molar alveoli; and orthodont incisors.
Etymology: Named for Kenneth Peter Aplin (1958–2019), for his contribution to Australian palaeontology and the taxonomy and systematics of Australasian murids.
Description
Skull. Two partial skulls are known (QMF52014 and 55753, Fig. 5A,B View Figure 5 ). The lacrimals, jugals, and much of the posterior of the skull and basicranium are missing from both specimens.
The nasals appear to be consistent in width along preserved length, tapering sharply at posterior contact with frontals.
Premaxilla short and robust. Anterior palatal foramen short, narrow, tapering abruptly at extremities, occupying similar area of premaxilla and maxilla. Anterior palatal foramen roughly half of length anterior of M 1. Narrow crest on ventral surface of maxilla between junction with premaxilla and anterior margin of M 1 variably developed, likely associated with age (some specimens, e.g., QMF55541, have it developed to an extreme degree, forming a blade. Fig. 5C View Figure 5 ). Zygomatic plate long, anterior edge straight, evenly curving posteriorly at dorsal end into zygomatic arch. Maxillary portion of zygomatic arch slopes posteroventrally at approximately 45° angle, almost reaching level of molar alveoli.
Palatine contacts maxilla level with posterior margin of M 1. Posterior palatal foramen level with M 2. Palate terminates in small, blunt postpalatal spine, approximately level with most posterior point of maxilla.
Frontals with sharp corner between temporal and orbital faces. Very small postorbital processes on squamosals, not associated with sutures. Supraorbital ridges not evident on frontals; parietal crests weakly developed on dorsal margin of squamosals and parietals. Braincase not greatly inflated, relatively flat dorsally, width exaggerated on QMF52014 by parting of parietals at midline.
I 1. Proportionally deep (I 1 depth/width of QMF52014 = 2.98/ 1.66 mm), orthodont. One paratype ( QMF55753 , Fig. 5B View Figure 5 ) appears to retain orange pigment in enamel, although this may instead be diagenetic iron staining .
M 1. Crown elongate, rounded anterior margin and angular posterior margin. Lophs sloped posteriorly. Accessory cusp and anterior cingulum absent. Lingual cusps bulge lingually at bases, giving lingual margin of crown an irregular appearance. Buccal cusps do not bulge at bases. T1 oval-shaped in occlusal outline, oriented antero-buccally postero-lingually. T1 postero-lingual of T2 . T1 separated from T2 by shallow cleft; T1 and T2 join after moderate wear. T2 broad and robust. T3 directly buccal of T2 , posterior margins of T2–3 form straight line. T3 small, fused to T2 . T3 discernible from T2 by shallow, poorly defined groove on anterior face of T2–3 complex. T4 subcircular in occlusal outline when unworn, becomes subtriangular after wear. In occlusal outline, T4 projects to a point anterior of the junction with T5 ; tapers posteriorly. T4 tapers towards T5 , separated by a shallow cleft, joined after moderate wear. T5 broad, boomerang-shaped in occlusal outline, with bulk of T5 antero-buccal of T4 and antero-lingual of T6 . T6 poorly defined, variably separated from T5 by shallow groove on anterior face of T5–6 complex. T6 broad, oriented antero-lingually postero-buccally, continuous with buccal half of T5 . T5–6 complex roughly parallel with T8–9 complex. T7 appears absent, although one specimen ( QMF55522 , Fig. 7C View Figure 7 ) has a bulge in the posterior loph that could be interpreted as a T7 fused to T8 . T8–9 complex broad, based between buccal margin of crown and posterior point of T4 . T9 fused to T8 , poorly defined by change in angle of anterior margin of occlusal surface of T8–9 complex. Very small posterior indent associated with posteroloph, commonly not visible in occlusal view .
Fine enamel ornament present on anterior faces of all lophs. M 1 has four roots: anterior, two lingual (commonly fused close to crown), and postero-buccal. Molar roots commonly split into multiple rootlets at tips. Alveoli of lingual roots variably fused, creating appearance of a single elongate lingual root.
M 2. Elongate, tapering posteriorly. Lingual cusps bulge lingually at bases, buccal cusps do not. T1 forms antero-lingual corner of crown. T1 subcircular in occlusal outline when slightly worn, becomes subtriangular (tapering buccally and posteriorly) after wear. T2–3 absent. Position of T3 variably marked by shallow depression on anterior face of T5–6 complex. T4 directly posterior of T1. T4–6 loph essentially identical to that on M 1. T8–9 complex based between buccal margin of T6 and posterior point of T4. T8–9 tapers slightly but does not form a point. T8 and T9 not differentiated. A ridge on the lingual side of T8 may represent a T7. Very small posterior indent associated with posteroloph, commonly not visible in occlusal view. Fine enamel ornament on anterior faces of both lophs, possibly less developed than that on M 1. M 2 has four main roots: antero-buccal, postero-buccal, postero-lingual, and antero-lingual. The antero-lingual and postero-lingual roots are variably joined. The antero-buccal root is variably bifurcated at the tip into two small rootlets.
M 3. Compact and simplified, moderately reduced. Some specimens (e.g., QMF55528, Fig. 7H View Figure 7 ) subcircular in occlusal outline. T1 well defined, rounded, oval-shaped in occlusal outline. T2–3 absent. Individual cusps of T4–6 loph not discernible. T4–6 loph gently curved, most anterior point at presumed location of T5. T4–6 loph sloped posteriorly. Posterior cusp broad, slightly narrower than T4–6 loph. Posterior cusp upright, very close to T4–6 loph. Some specimens (e.g., QMF55530, Fig. 7G View Figure 7 ) have posterior cusp very close to “T4” but larger gap separating posterior cusp from “T6”. Posterior cusp oval-shaped in occlusal outline. One specimen (QMF55544) has a small posterior cingulum cusp. M 3 has four roots: antero-buccal, posterior, and joined antero-lingual and lingual.
Mandible. No specimens are completely intact, with all displaying degrees of damage to the posterior processes and incisor alveolus. Mandible deep and robust, with deepest point ventral of M 1. M 1 longer than M 2, but similar width. M 3 smaller than M 2, but not heavily reduced. Coronoid process damaged or missing on all specimens, but appears to be taller than articular process. Articular process projects slightly posterior of angular process. Angular process damaged on all specimens, but appears to be rounded. Mental foramen ventral of dorsal inflection of diastema. Superior masseteric crest very poorly defined below molars; inferior masseteric crest well developed, terminates anteriorly posterior of mental foramen. Mandibular symphysis marked by dorsal crest in anterior part of diastema; symphysis ends ventrally of anterior root of M 1. Incisor alveolus forms prominent tubercle on buccal surface of ascending ramus, although this is damaged in all specimens. Postalveolar ridge sharply defined below and posterior of M 3, less defined posterior of retromandibular fossa. Retromandibular fossa small in young adult individuals, greatly expanded in mature individuals (assessed on the basis of molar wear).
I 1. Proportionally deep (I 1 depth/width of QMF55542 [subadult] = 1.98/1.13, adults proportionally deeper). One specimen (QMF55543, Fig. 6B View Figure 6 ) may retain orange pigment in enamel, but lost in majority of specimens.
M 1. Rounded anteriorly, subrectangular posteriorly. Anterior and middle lophids crowded together. Anterior lophid upright, middle, and posterior lophids sloped anteriorly. Anterior lophid narrower than middle lophid. Middle and posterior lophids of similar width. Antero-buccal cuspid small, subcircular in occlusal outline, fused to antero-lingual cuspid. Antero-buccal and antero-lingual cuspids only distinguishable when unworn, form single anterior lophid after moderate wear. Antero-lingual cuspid much larger than antero-buccal cuspid, forms much of the anterior lophid. Unworn specimens show antero-lingual cuspid with two buccal extensions: one joining the antero-buccal cuspid, the other directly posterior and postero-lingual of the antero-buccal cuspid between the main body of the anterior lophid and the middle lophid. Antero-buccal cuspid separated from protoconid by shallow cleft, eliminated by wear on some specimens; antero-lingual cuspid separated from metaconid by relatively deeper cleft, more resistant to wear.
Protoconid subtriangular in occlusal outline, tapering lingually to join metaconid and posteriorly along buccal margin of crown. Metaconid subequal in size to protoconid. Metaconid subtriangular in occlusal outline, tapering buccally to join protoconid, tapering slightly posteriorly and anteriorly. Anterior margin of middle lophid buccally perpendicular to long axis of crown, curves antero-lingually to most anterior point of metaconid. Posterior face of middle lophid curved, bowing anteriorly between most posterior points of protoconid and metaconid.
Entoconid directly posterior of metaconid. Entoconid subtriangular in occlusal outline, tapering buccally to join hypoconid and posteriorly to a lesser degree. Hypoconid directly posterior of protoconid, buccal and slightly posterior of entoconid. Hypoconid subtriangular in occlusal outline, tapering lingually to join entoconid, and posteriorly to a lesser degree. Hypoconid and entoconid variably have small anterior extensions. Hypoconid projects slightly further posteriorly than entoconid.Anterior edge of occlusal surface of posterior lophid commonly straight, but some specimens (e.g., QMF55533, Fig. 7K View Figure 7 ) have a slight bulge, at approximately the midline of the crown. Posterior margin of occlusal surface curved, bowed anteriorly with most anterior point directly posterior of midline junction between hypoconid and entoconid. Posteroconid tolerably well developed, lenticular in occlusal outline, bound by bases of hypoconid and entoconid. Posteroconid does not project beyond posterior margin of crown. Fine enamel ornament on posterior faces of middle and posterior lophids, not visible on anterior lophid due to close proximity of middle lophid. M 1 has three roots: anterior, a broad posterior, and a small lingual root.
M 2. Crown roughly square in occlusal outline, with rounded corners. Both lophids sloped anteriorly. Protoconid larger than metaconid, both at apex and base. Protoconid tear-shaped in occlusal outline, tapering lingually to join metaconid at midline of crown. Metaconid directly lingual of protoconid, tear-shaped in occlusal outline, tapering buccally. Unworn specimens (e.g., QMF55536, Fig. 7N View Figure 7 ) have no cleft separating protoconid and metaconid. Hypoconid directly posterior of protoconid. Hypoconid tear-shaped when unworn, becomes subtriangular after light wear. Hypoconid tapers antero-lingually to join entoconid at midline of crown. Hypoconid tapers posteriorly further than entoconid. Entoconid slightly less robust than hypoconid. Entoconid tear-shaped in occlusal outline, tapering directly buccally, meeting hypoconid at an angle. No separation between hypoconid and entoconid.
Posteroconid well developed, lenticular in occlusal outline. Posteroconid commonly centred on midline of crown, although one specimen ( QMF55535 , Fig. 7 O View Figure 7 ) has it centred slightly buccal of the midline. Posteroconid projects slightly beyond posterior margin of crown. Fine enamel ornament on posterior faces of lophids. M 2 has two broad roots: anterior and posterior .
M 3. Almost triangular in occlusal outline, with heavily rounded corners. Protoconid slightly larger than metaconid. Protoconid tear-shaped in occlusal outline, tapering lingually to join metaconid. Metaconid tear-shaped in occlusal outline, tapering buccally to join protoconid. Protoconid and metaconid joined by narrow ridge. Posterior lophid broad, commonly supplemented by small cuspid on buccal side. Posterior lophid shaped like an elongate oval in occlusal outline, supplementary cuspid subcircular. Supplementary buccal cuspid variably separated from posterior lophid by shallow cleft or fused. M 3 has three roots: posterior, and fused antero-buccal and antero-lingual.
Remarks
Uromys aplini can be distinguished from other members of Uromys (Uromys) as follows: Uromys aplini differs from U. caudimaculatus by being smaller; having a less elongate rostrum; having a smaller posterior indent in T8–9 on M 1–2; and having shorter anterior palatal foramina. Uromys aplini differs from U. sherrini by being smaller; having a less elongate rostrum; and having a more reduced M 3 / 3. Uromys aplini differs from U. hadrourus by being larger; having proportionally shorter anterior palatal foramina; having a proportionally shorter rostrum; by commonly possessing a crest on the maxilla between the maxilla/premaxilla contact and M 1; and having a zygomatic arch that plunges further ventrally, reaching the level of the molar alveoli. The molars of Uromys aplini could not be effectively compared to those of U. hadrourus , as all examined specimens of the latter were heavily worn. Uromys aplini differs from U. anak by being smaller; having a less elongate rostrum; lacking postorbital processes; having a smaller posterior indent in T8–9 on M 1–2; and having the anterior palatal foramina shared equally between the premaxilla and maxilla. Uromys aplini differs from U. neobrittanicus by being smaller; lacking large postorbital processes; having the skull relatively flat dorsally; and having parietals that are roughly rectangular in dorsal outline. Uromys aplini differs from U. emmae by being smaller; having a deeper zygomatic arch; having the zygomatic plate not projecting as far anterior of the zygomatic arch; and having the anterior palatal foramina shared equally between the premaxilla and maxilla. Uromys aplini differs from U. boeadii by being smaller; having smaller postorbital processes; and lacking supraorbital ridges.
Only one skull of U. siebersi is known, and this specimen was not available for the current study. But a measurement of the molar row (13.3 mm) provided by Groves & Flannery (1994) shows that U. siebersi is larger than U. aplini in this aspect ( Table 2). Thomas (1923b) also provided measurements, though these are less precise than currently obtainable with modern precision measuring tools. The interorbital width and length of the “palatal foramina” (presumably the anterior palatal foramina) are both larger (10.3 mm and 7 mm, respectively, versus 8.34 mm and 6.00 mm for U. aplini ).
Uromys aplini is hitherto known mostly from deposits at Mount Etna that are dominated by taxa that had ecological affinities to rainforest environments. The oldest deposits that yield the species are>500 ka, whilst the youngest is 205–170 ka.
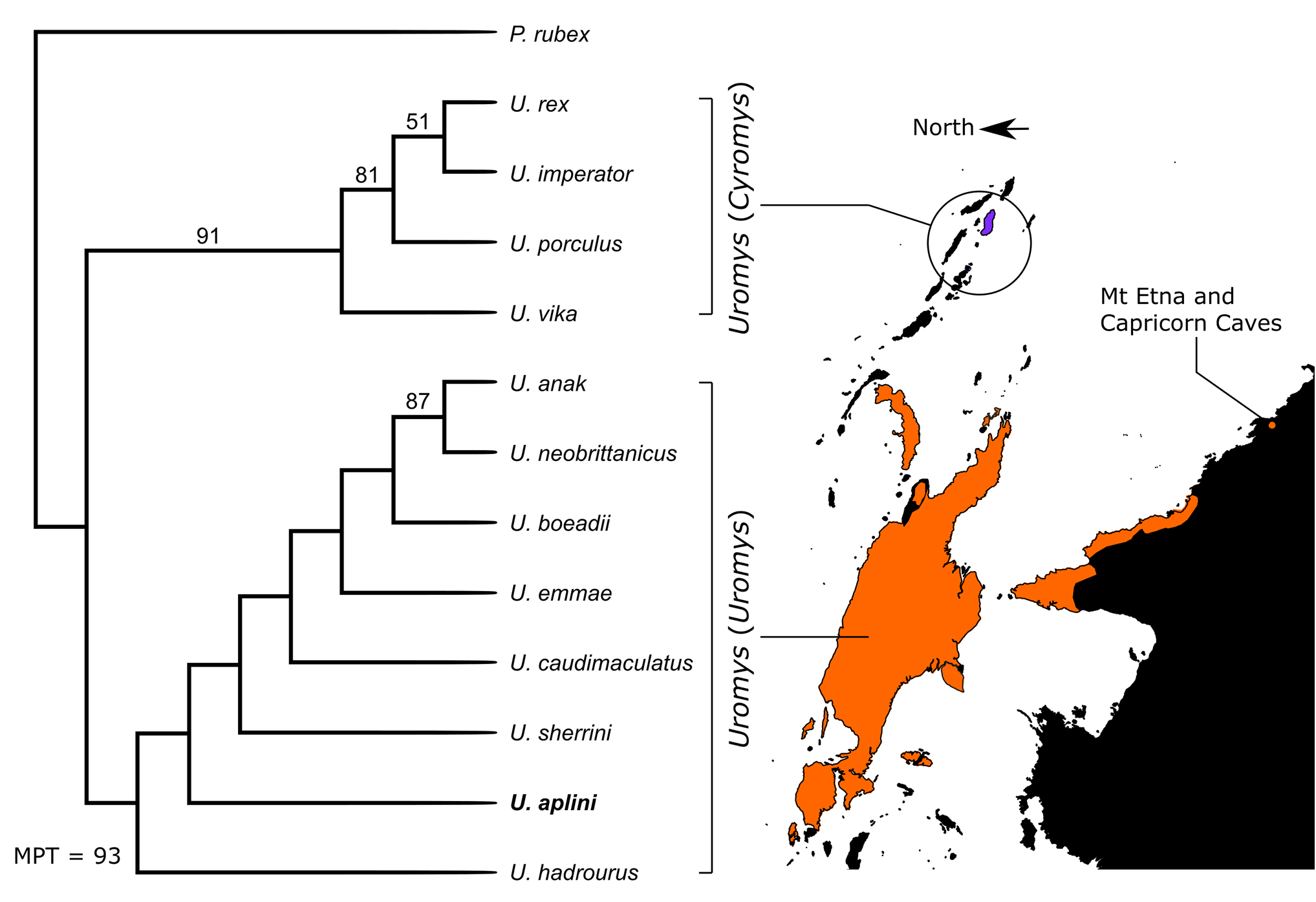
Phylogenetic analysis
Our phylogenetic analysis returned topological features similar to that recovered by Groves & Flannery (1994). We used Paramelomys rubex as the most appropriate outgroup taxon to polarize the character-states within Uromys . Thirtythree characters were parsimony informative with seven uninformative and considered to be autapomorphies of these taxa. The derived character states for characters 12, 24 and 25 are considered to be autapomorphies of U. rex , so are uninformative in relation to U. aplini . The derived character states for characters 34 and 38 are considered to be autapomorphies of U. imperator and U. emmae respectively, also uninformative for the fossil taxon. Finally, uninformative characters 15 and 37 are restricted to U. hadrourus , with the derived state of character 15 an autapomorphy and 37 ambiguous due to the missing states in the fossil taxon ( U. aplini ) and in U. sherrini .
The parsimony analysis returned two most parsimonious trees (MPT) of 93 steps ( Fig. 9 View Figure 9 ). Both MPTs consistently returned a basal split with one clade solely composed of species within the subgenus Cyromys and found today in the Solomon Islands group ( U. imperator , U. rex , U. porculus , and U. vika ). The Cyromys clade is strongly supported by bootstrap value of 91%. The other clade is composed solely of species within the subgenus Uromys and includes our fossil taxon, U. aplini . Although this clade is poorly supported, it is likely that the large amount of missing data and morphological variability of U. caudimaculatus have created internal instability within this clade. Further characterization of U. caudimaculatus subspecies and better resolution of missing data may increase the support for the monophyly of Uromys (Uromys) and Uromys (Cyromys) .
Resolution within the Uromys (Uromys) clade is poor, although the New Guinean U. anak and New Britain U. neobrittanicus are strongly supported (87%) as sister taxa. The positions of the remaining taxa are poorly supported by bootstrap values, but both MPTs return identical positions of all species, suggesting that the overall topology is valid. At the base of the clade lie the Australian Uromys hadrourus , the fossil taxon U. aplini , and U. sherrini . In a more derived position, sister to these Australian endemic species, is U. caudimaculatus , which is then sister taxon to a clade containing the northern New Guinean island endemics ( U. emmae and U. boeadii ), the mainland New Guinea U. anak , and U. neobrittanicus from New Britain.
The two basal clades, comprising Uromys (Cyromys) and Uromys (Uromys) , were supported by Groves & Flannery (1994), so this result is not surprising. But our analysis, using Paramelomys as the outgroup, suggests that the Australian Uromys are basal to the Uromys (Uromys) clade.
The Middle Pleistocene age of our phylogenetically basal extinct taxon ( U. aplini ) is younger than the divergence time estimates (e.g., Early Pleistocene) for the more derived extant species within the clade (Watts & Baverstock, 1994; Bryant et al., 2011). This would probably preclude U. aplini from being a chronospecies of the extant Australian species of Uromys (Uromys) .

| LU |
St. Petersburg University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.